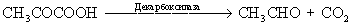
Углеводный обмен, процессы усвоения углеводов в организме; их расщепление с образованием промежуточных и конечных продуктов (деградация, диссимиляция), а также новообразование из соединений, не являющихся углеводами (глюконсогенез), или превращение простых углеводов в более сложные. Под влиянием пищеварительных ферментов гидролаз (различного типа амилаз, гликозидаз) сложные поли- и олигосахариды подвергаются расщеплению до моносахаридов — гексоз или пентоз, которые утилизируются организмом. Полисахариды ферментативно расщепляются также фосфорилазами с образованием глюкозо-1-фосфата. Деградация гексоз, поступивших в клетку, осуществляется в процессе брожения или гликолиза, а также окислением в пентозофосфатиом цикле.
Брожение и гликолиз представляют собой анаэробные (без участия кислорода воздуха) пути деградации моносахаридов, завершающиеся при брожении образованием этилового спирта, высших спиртов, масляной или пропионовой кислот, а при гликолизе и молочнокислом брожении — образованием молочной кислоты. Начальной реакцией, обязательной для последующих превращений моносахаридов, является их фосфорилирование, катализируемое ферментом гексокиназой. При анаэробном процессе на следующем этапе происходит повторное фосфорилирование, завершающееся образованием дифосфорного эфира фруктозы, который расщепляется альдолазой на две фосфотриозы (завершение 1-й стадии брожения или гликолиза). В дальнейшем в результате окислительно-восстановительных реакций последовательно образуются фосфоглицериновые кислоты и фосфоенолпиро-виноградная кислота. Эти реакции сопровождаются связыванием минерального фосфата, переносом остатка фосфорной кислоты на аденозиндифосфат (АДФ) и образованием аденозинтрифосфата (АТФ). Совокупность этих реакций составляет 2-ю стадию анаэробных превращений углеводов, играющих существенную роль в образовании богатых энергией фосфорных соединений (см. Макроэргические соединения).
Различие между спиртовым брожением, с одной стороны, и гликолизом или молочнокислым брожением — с другой, выявляется на стадии превращения пировиноградной кислоты (пирувата): при спиртовом брожении в клетках под влиянием пируватдекарбоксилазы образуются CO2 и уксусный альдегид, восстанавливаемый алкогольдегидрогеназой в спирт:
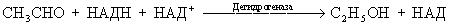
При гликолизе или молочнокислом брожении пируват не подвергается анаэробному декарбоксилированию, а восстанавливается в молочную кислоту лактатдегидрогеназой:  Пируватдегидрогеназа, представленная в животных тканях и микроорганизмах, осуществляет декарбоксилирование пирувата с использованием кислорода и образованием ацетилкофермента А (ацетил-КоА), вовлекая таким образом пируват в цикл трикарбоновых кислот (см. Трикарбоновых кислот цикл). Полное окисление пирувата происходит в результате троекратного декарбоксилирования и пятикратной дегидрогенизации в цикле трикарбоновых кислот: CH3CO. COOH +
Пируватдегидрогеназа, представленная в животных тканях и микроорганизмах, осуществляет декарбоксилирование пирувата с использованием кислорода и образованием ацетилкофермента А (ацетил-КоА), вовлекая таким образом пируват в цикл трикарбоновых кислот (см. Трикарбоновых кислот цикл). Полное окисление пирувата происходит в результате троекратного декарбоксилирования и пятикратной дегидрогенизации в цикле трикарбоновых кислот: CH3CO. COOH + 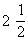 O2 ® 3CO2 + 2H2O. Этот процесс выходит за рамки У. о., однако может рассматриваться как завершающая его стадия: окисление продукта гликолиза — пирувата.
O2 ® 3CO2 + 2H2O. Этот процесс выходит за рамки У. о., однако может рассматриваться как завершающая его стадия: окисление продукта гликолиза — пирувата.
Окислительное превращения углеводов (пентозный путь, или пентозофосфатный цикл) также начинаются с глюкозомоно-фосфата. Затем происходят последовательно 2 дегидрогеназные реакции: первая приводит к фосфоглюконовой кислоте, а вторая — к освобождению CO2 и образованию фосфопентозы. Важным итогом этих окислительных реакций является образование восстановленного никотинамидадениндинуклеотидфосфата — кофермента, участвующего во многих синтезах (например, в синтезе жирных кислот). Последующие реакции пентозного пути не связаны с использованием молекулярного кислорода и протекают в анаэробных условиях. При этом частично образуются вещества, характерные для 1-й стадии гликолиза (фруктозо-6-фосфат, фруктозодифосфат, фосфотриозы), а частично специфические для пентозного пути (седогептулозо-1-фосфат, седогептулозо-1,7-дифосфат, фосфопентозы, фосфотетроза, а, возможно, также фосфорные эфиры моносахаридов с 8 атомами углерода). Перечисленные вещества, характерные для гликолиза и пентозного пути, могут участвовать в обратимых реакциях взаимопревращения. Аналогичные реакции протекают и при фотосинтезе на стадиях образования фосфопентоз из фруктозо-6-фосфата и фосфотриозы (так называемый цикл Кальвина).
Пути биосинтеза углеводов представлены в живых клетках процессами глюконеогенеза и образованием высокомолекулярных полисахаридов. Процесс глюконеогенеза начинается с карбоксилирования пирувата при участии сложной по своей структуре ферментной системы пируваткарбоксилазы, приводящей к образованию щавелевоуксусной кислоты (ЩУК) с участием в качестве кофермента биотина. Стимулирует эту реакцию ацетил-КоЛ. В свою очередь, ЩУК подвергается в цитоплазме реакции декарбоксилирования под действием фермента фосфоенолпируваткарбоксикиназы. Благодаря этим реакциям преодолеваются энергетические барьеры и может образоваться из пирувата фосфоенолпировиноградная кислота — источник глюкозы. В фотосинтезирующих бактериях реализуется также и др. возможность: обращение цикла трикарбоновых кислот, восстановление при этом 3 молекул CO2 и образование фосфоенолпирувата. У растений и микроорганизмов в процессе глюконеогенеза важную роль играет глиоксилатный цикл.
Суммарное уравнение реакций, ведущих от пирувата к глюкозе, может быть записано следующим образом:
2CH3COCOOH + 4АТФ + 2ГТФ + 2НАДН + 2H + 6H2O ® глюкоза + 2НАД + 4АДФ + 2ГДФ + 6 неорганический фосфат
(где АТФ — аденозинтрифосфат, а ГТФ — гуанозинтрифосфат). Синтез олиго- и полисахаридов при участии различных гликозилтрансфераз осуществляется путём переноса гликозильного остатка с нуклеозиддифосфатсахара на моносахарид или же на концевой остаток моносахарида в молекуле поли- или олигосахарида. Таким образом, цепь, состоящая из гексозных остатков, удлиняется. Ветвление амилопектина или гликогена за счёт образования 1,6-связей осуществляется ферментом амило- (1,4—1,6) — трансгликозилазой, катализирующим перенос концевого фрагмента, состоящего из 6 или 7 гликозильных остатков, с конца главной цепи на гидроксильную группу 6-го углеродного атома остатка глюкозы какой-либо из цепей полисахарида.
Пути регуляции У. о. крайне разнообразны. На любых уровнях организации живого У. о. регулируется факторами, влияющими на активность ферментов, участвующих в реакциях У. о.: концентрацией субстратов и продуктов отдельных реакций, кислородным режимом, температурой, проницаемостью биологических мембран, определяющей возможность контакта между участниками реакций, концентрацией коферментов, необходимых для отдельных реакций, и т.д. У животных на всех стадиях синтеза и распада углеводов регуляция У. о. осуществляется с участием нервной системы и гормонов.
Лит.: Кретович В. Л., Основы биохимии растений, 5 изд., М., 1971; Шлегель Г., Общая микробиология, [пер. с нем.], М., 1972; Ленинджер А., Биохимия, пер. с англ., М., 1974. См. также ст. Обмен веществ и литературу при ней.
С. Е. Северин.